|
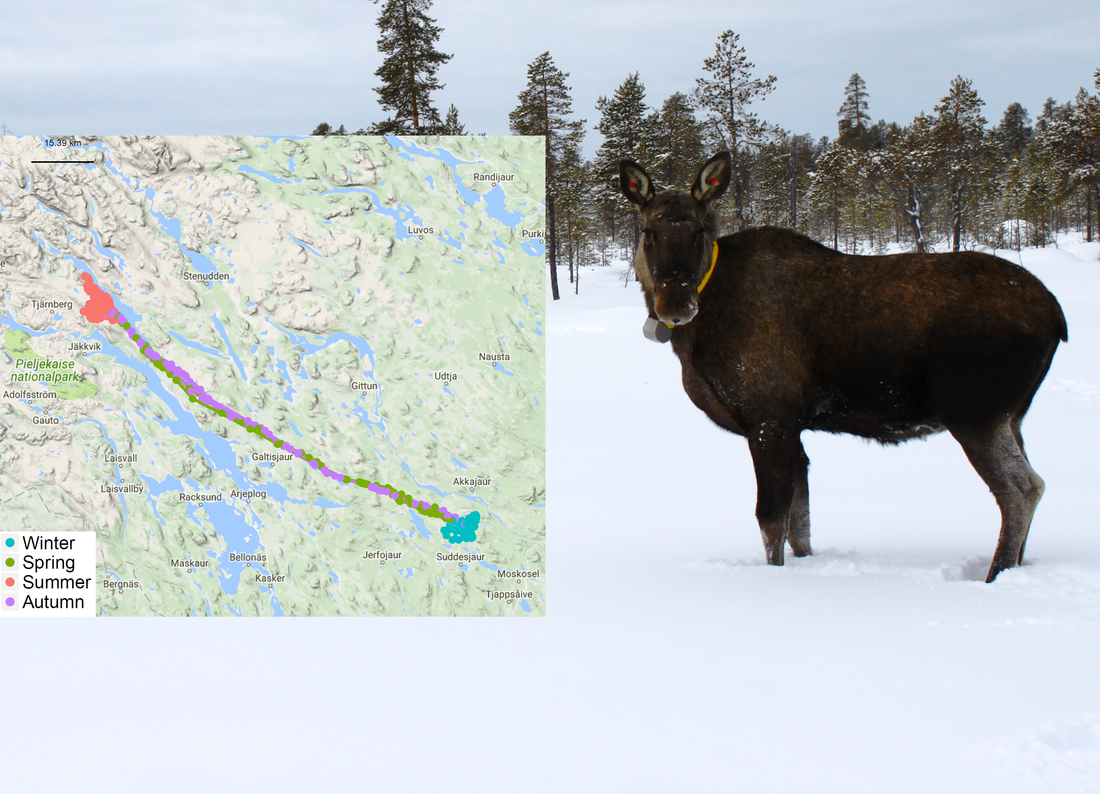
Linking movement ecology to wildlife management: Managing moose in SwedenHere is a brief overview of my PhD project conducted between Nov'12 and Nov'16. As I write this, four articles have been accepted for publication, a fifth has been submitted and the sixth is still in progress. I have written an extensive overview of my thesis work in a summarising chapter, called a "kappa" in Swedish, which is available here: http://pub.epsilon.slu.se/13745/1/Allen_A_161024.pdf. Below is a shorter summary of each of the papers from my thesis. Each picture can be enlarged when you click on it. If you have any questions about my research, please do not hesitate to contact me.
|
Paper I - Movement-Management Framework
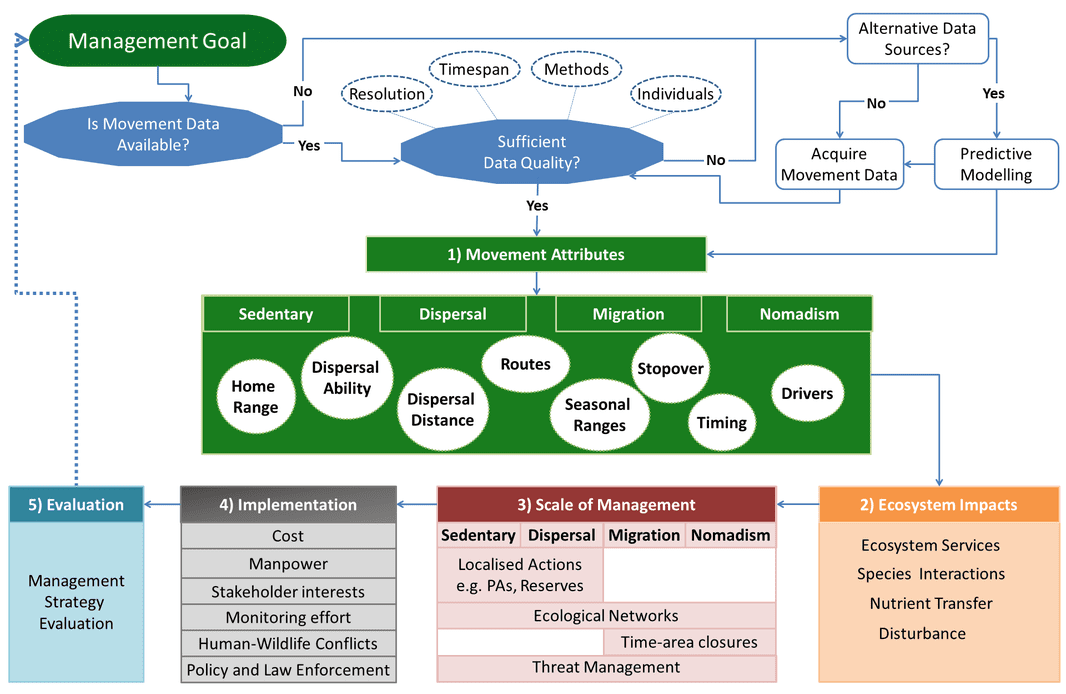
The movement-management framework
|
The first paper in my thesis, published in Frontiers in Ecology and Evolution, reviews the opportunities for linking movement ecology to wildlife management, and also why we need to incorporate movement into management. We present a conceptual movement-management framework that consists of five steps where you first identify the movements of a species and how it impacts an ecosystem in order to define the scale of management. The scale of management is linked to the implementability (e.g. feasability) of management actions and the final steps consists of evaluation to create an iterative adapative management cycle. We illustrate the framework in a case study using Baltic Salmon in the paper, whilst I have used the framework to structure my research questions and I also apply our framework to managing moose in Sweden. The full paper is freely available here.
|
Paper II - Classifying and Quantifying movements using NSD
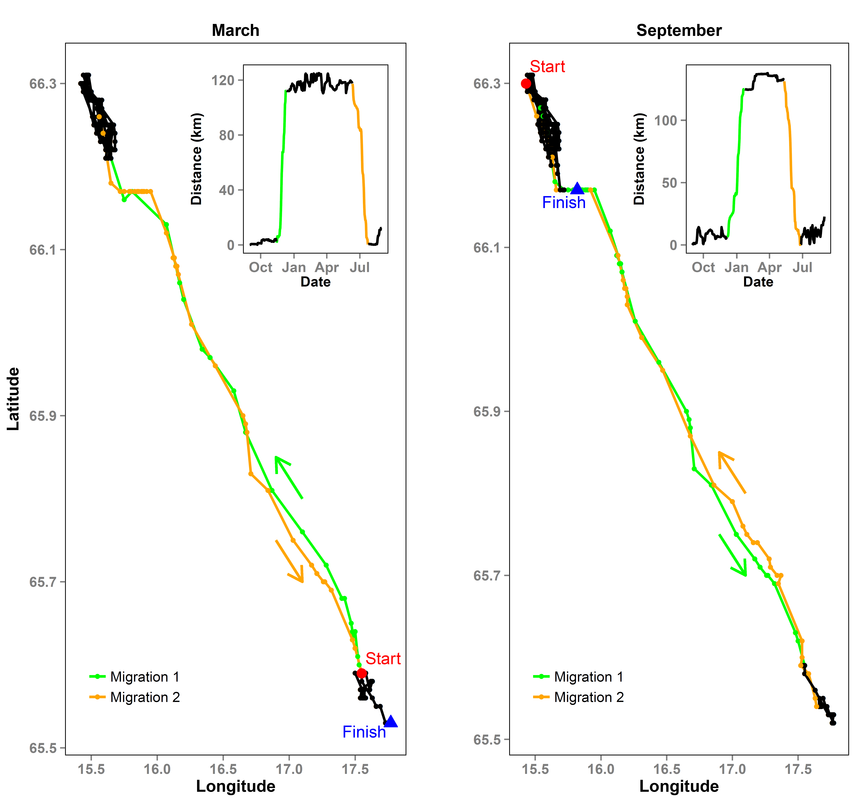
Example of spring and autumn migrations and the associated NSD
|
The second paper in my thesis, published in PLoS ONE, provides guidance for using the Net-Squared Displacement (NSD) method. The NSD method is a model-driven approach for firstly classifying movements as sedentary, (mixed) migratory, dispersing or nomadic, and then quantifying these movements by extracting the distance of movements, their timing and duration. A number of challenges have been described when using the method, and we aim to overcome these challenges by providing advice regarding certain aspects of the method. We do this by addressing three questions related to the scale of species movement, the starting location (in time and space) and the resolution of the data. We first classify movements of 319 moose, and then explore issues in quantifying movements using 41 migratory individuals tracked at the same time. We recommend users to classify movements using the mean-squared displacement (MSD), and then to quantify movements using the NSD. We found that one location a day is sufficient for classifing annual movements, and adding more locations per day may actually reduce the performance of the method. A mean starting location (e.g. a mean location during the first month) improves classification as the location is more likely to be near the centre of the home range. Initial exploratory analyses are recommended to ensure that the starting date is not during a period of migration (for e.g. during May/Jun for spring migrations of moose). The full paper is freely available here.
|
Paper III - Understanding intraspecific variation in space use
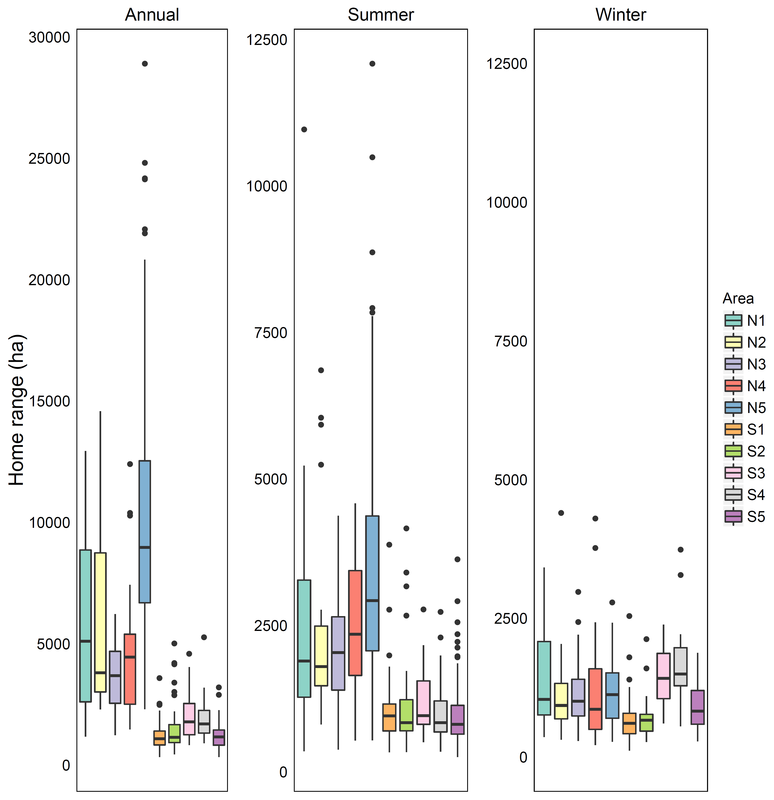
Variation in home range size for the ten study populations in the paper
|
Individuals exhibit large variation in their movement patterns and much research has aimed at understanding what causes this variation. This knowledge is important for understanding what environmental and life history factors may shape an individual's movement, and thus improve our ability to derive estimates of a population mean. This may guide the size of management areas, or determine the best timing of management actions based on for example the average time that individuals arrive at and depart from their summer ranges. We analysed intraspecific variation in movement patterns of over 500 annual trajectores from 307 moose than occur in ten different populations. This allowed us to analyse variation at two scales that include variation among popualtions and variation within each popualtion. We used the NSD approach to classify and quantify movements, and used this information to derive seasonal home ranges. We then used multiple environmental variables and life history data (age, sex) to determine which factors best explained variation among individuals. We identified variation in HRs across a broad scale with factors related to the climate explaining most variation in HR size. In contrast, variation among individuals within the same population was explained by the life history of the individual and factors related to the landscape such as the proportion of preferred habitats or the structure of the landscape. Our paper provides insights to how intraspecific variation in HR size is due to factors operating and both broad and fine scales, and it is important to understand the influence of these factors when identifying average movements of a population. This work has been published in Ecosphere and is freely available here.
|
Paper IV - Habitat-performance relationships
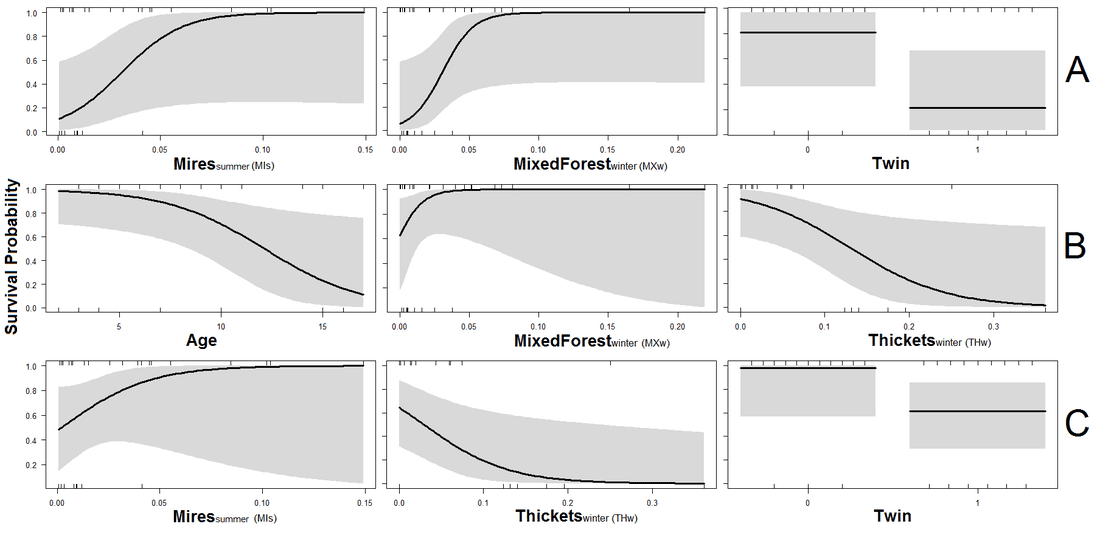
The top three models that explain variation in calf survival. Females that spent more time in Thicket type habitats in winter had lower calf survival, as did older females, whilst females that spent more time in Mire type habitats in summer had higher calf survival
|
A risk in the field of movement ecology is that researchers only get excited about the movements they observe without considering what they mean for the ecology of the species. In the fourth paper of my thesis, we investigate how the space use patterns of females affect reproduction and calf survival to understand why an island population of moose are declining. We monitored 17 females using GPS collars over three years and also recorded whether they had calves, and if the calves survived. Calving and fecundity rates of females were comparable to other Scandinavian popualtions of moose and thus reproduction does not appear to be an obvious cause for the decline. In contrast, calf survival was often lower than 25%. No predators of moose occur on Öland and calf survival is usually greater than 70% in most other Swedish populations. Understanding the causes of low calf survival therefore appears to be vital for determining why the population is declining. We observed clear habitat-performance relationships. Females that spent more time in thicket-type habitats during winter had lower calf survival the following spring. In contrast, time spent in mixed forest during winter, or mire habitats during summer, resulted in higher calf survival. Our results provide managers and researchers with some indications of factors influencing the performance of moose. We also discuss how climate change may be affecting moose and potential future impacts. The work has been published in Ecology and Evolution and is freely available here.
|
